



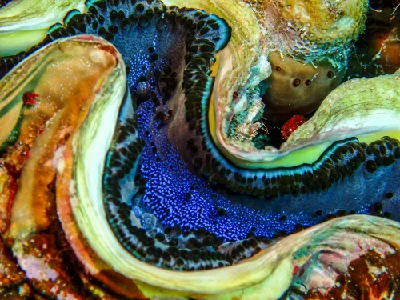


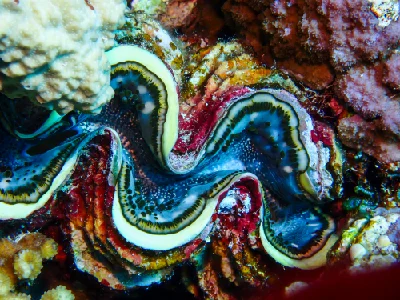



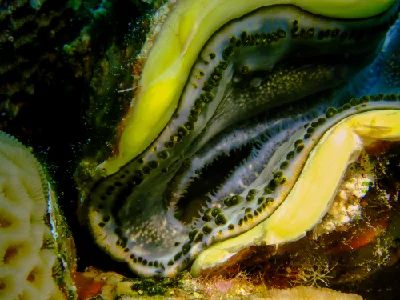
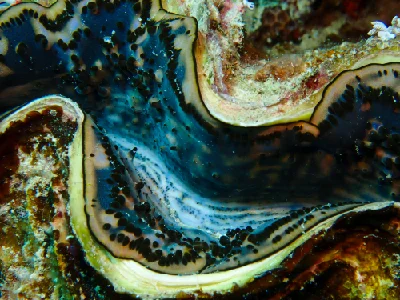




The giant clams are the members of the clam genus Tridacna that are the largest living bivalve molluscs. There are several species of "giant clams" in the genus Tridacna, which are often misidentified for Tridacna gigas, the most commonly intended species referred to as "the giant clam".
Tridacna gigas is one of the most endangered clam species. Antonio Pigafetta documented these in his journal as early as 1521. One of a number of large clam species native to the shallow coral reefs of the South Pacific and undefineds, they can weigh more than 200 kilograms (440 lb), measure as much as 120 cm (47 in) across and have an average lifespan in the wild of over 100 years. They are also found off the shores of the 🇵🇭 Philippines and in the South China Sea in the coral reefs of Sabah (🇲🇾 Malaysian Borneo).
The giant clam lives in flat coral sand or broken coral and can be found at depths of as much as 20 m (66 ft). Its range covers the Indo-Pacific, but populations are diminishing quickly, and the giant clam has become extinct in many areas where it was once common. The maxima clam has the largest geographical distribution among giant clam species; it can be found off high- or low-elevation islands, in lagoons or fringing reefs. Its rapid growth rate is likely due to its ability to cultivate algae in its body tissue. Although larval clams are planktonic, they become sessile in adulthood. The creature's mantle tissues act as a habitat for the symbiotic single-celled dinoflagellate algae (zooxanthellae) from which the adult clams get most of their nutrition. By day, the clam opens its shell and extends its mantle tissue so that the algae receive the sunlight they need to photosynthesize.
Young Tridacna gigas are difficult to distinguish from other species of Tridacninae. Adult Tridacna gigas are the only giant clams unable to close their shells completely. Even when closed, part of the mantle is visible, unlike the very similar Tridacna. derasa. However, this can only be recognized with increasing age and growth. Small gaps always remain between shells through which retracted brownish-yellow mantle can be seen. Tridacna gigas has four or five vertical folds in its shell; this is the main characteristic that separates it from the similar shell of Tridacna derasa, which has six or seven vertical folds. As with massive deposition of coral matrices composed of calcium carbonate, the bivalves containing zooxanthellae have a tendency to grow massive calcium carbonate shells. The mantle's edges are packed with symbiotic zooxanthellae that presumably utilize carbon dioxide, phosphates, and nitrates supplied by the clam. The mantle border itself is covered in several hundred eyespots about 0.5mm in diameter. Each one consists of a small cavity containing a pupil-like aperture and a base of one hundred or more photoreceptors. These receptors allow Tridacna gigas to respond to sudden dimming of light by withdrawing their mantles and partially closing their shells, presumably to protect from potential predators. They do not retract their mantles in response to increased illumination, but it has been observed that a change in the direction of light results in a shift in mantle orientation. In addition to a dimming response, Tridacna gigas also responds to the movement of an object before a shadow has been cast. In order for this to happen, an image forming optical system is required as the response is based on the local dimming of one part of the generated image relative to the rest. This sequential dimming of receptors caused by the movement of a dark object allows enough time for the mantle to be retracted before a potential predator is directly overhead and casting a shadow.
The largest known Tridacna gigas specimen measured 137 centimeters (4 ft 6 in). It was discovered around 1817 on the northwestern coast of Sumatra (🇮🇩 Indonesia), 🇮🇩 Indonesia, and its shells are now on display in a museum in Northern Ireland. The joint weight of the two shells is 230 kilograms (510 lb), which suggests that the live weight of the animal would have been roughly 250 kilograms (550 lb). Another unusually large giant clam was found in 1956 off the 🇯🇵 Japanese island of Ishigaki. However, it was not examined scientifically before 1984. The shell's length was 115 centimeters (3 ft 9 in) and the weight of the shells and soft parts was 333 kilograms (734 lb). Scientists estimated the live weight to be around 340 kilograms (750 lb).
Giant clams are filter-feeders, yet 65-70 percent of their nutritional needs are covered by symbiotic unicellular algae (zooxanthellae). This enables giant clams to grow as large as one meter in length even in nutrient-poor coral-reef waters. The clams cultivate algae in a special circulatory system which enables them to keep a substantially higher number of symbionts per unit of volume.
Tridacna gigas reproduce sexually and are hermaphrodites (producing both eggs and sperm). Self-fertilization is not possible, but this characteristic does allow them to reproduce with any other member of the species. This reduces the burden of finding a compatible mate, while simultaneously doubling the number of offspring produced by the process. As with all other forms of sexual reproduction, hermaphroditism ensures that new gene combinations be passed to further generations. Since giant clams cannot move themselves, they adopt broadcast spawning, releasing sperm and eggs into the water. A transmitter substance called spawning induced substance (SIS) helps synchronize the release of sperm and eggs to ensure fertilization. The substance is released through a syphonal outlet. Other clams can detect SIS immediately. Incoming water passes chemoreceptors situated close to the incurrent syphon, which transmit the information directly to the cerebral ganglia, a simple form of brain. Detection of SIS stimulates the giant clam to swell its mantle in the central region and to contract its adductor muscle. Each clam then fills its water chambers and closes the incurrent syphon. The shell contracts vigorously with the adductor's help, so the excurrent chamber's contents flows through the excurrent syphon. After a few contractions containing only water, eggs and sperm appear in the excurrent chamber and then pass through the excurrent syphon into the water. Female eggs have a diameter of 100 micrometres (0.0039 in). Egg release initiates the reproductive process. An adult Tridacna gigas can release more than 500 million eggs at a time. Spawning seems to coincide with incoming tides near the second (full), third, and fourth (new) quarters of the moon phase. Spawning contractions occur every two or three minutes, with intense spawning ranging from thirty minutes to two and a half hours. Clams that do not respond to the spawning of neighboring clams may be reproductively inactive.
The fertilized egg floats in the sea for about 12 hours until eventually a larva (trochophore) hatches. It then starts to produce a calcium carbonate shell. Two days after fertilization it measures 160 micrometres (0.0063 in). Soon it develops a "foot," which is used to move on the ground; it can also swim to search for appropriate habitat. At roughly one week of age, the clam settles on the ground, although it changes location frequently within the first few weeks. The larva does not yet have symbiotic algae, so it depends completely on plankton. Free floating zooxanthellae are also captured while filtering food. Eventually the front adductor muscle disappears and the rear muscle moves into the clam's center. Many small clams die at this stage. The clam is considered a juvenile when it reaches a length of 20 cm (8 in). It is difficult to observe the growth rate of Tridacna gigas in the wild, but laboratory-reared giant clams have been observed to grow 12 cm (4.7 in) a year. The ability for Tridacna to grow to such large sizes with fleshy mantles that extend beyond the edges of their shells is considered to be the result of total reorganization of bivalve development and morphology. Two evolutionary explanations were historically suggested for this process. Sir Yonge suggested and maintained for many years that the visceral-pedal ganglia complex rotate 180 degrees relative to the shell, requiring that they develop and evolve independently. Stasek proposed instead that the growth occurs primarily in a posterior direction instead of the more typical direction of ventral in
Why it's threatened
Threat classification from the IUCN Red List.



